węzeł zatokowo-przedsionkowy
węzeł zatokowo-przedsionkowy (SA) jest wrzecionowatą strukturą złożoną z macierzy tkanki włóknistej z ściśle upakowanymi komórkami. Ma 10-20 mm długości, 2-3 mm szerokości i grubości, zwężając się ku dolnej żyle żylnej (IVC). Węzeł SA znajduje się mniej niż 1 mm od powierzchni epikardialnej, bocznie w prawym przedsionku sulcus terminalis na skrzyżowaniu przednio-przyśrodkowego aspektu żyły głównej górnej (SVC) i prawego przedsionka (RA).,
tętnica zaopatrująca węzeł zatokowy odgałęzia się od prawej tętnicy wieńcowej w 55-60% serc lub od lewej tętnicy obwodowej w 40-45% serc. Tętnica zbliża się do węzła z kierunku zgodnego z ruchem wskazówek zegara lub przeciwnie do ruchu wskazówek zegara wokół węzła SVC-RA.
węzeł SA jest gęsto unerwiony postganglionicznymi terminalami nerwowymi adrenergicznymi i cholinergicznymi. Neuroprzekaźniki modulują szybkość rozładowania węzłów SA poprzez stymulację receptorów beta-adrenergicznych i muskarynowych. Zarówno podtypy adrenoceptorów beta1, jak i beta2 występują w węźle SA., Ludzki węzeł SA zawiera ponad 3-krotnie większą gęstość receptorów beta-adrenergicznych i muskarynowych cholinergicznych niż sąsiadująca tkanka przedsionkowa.
przewodnictwo Śródnabłonkowe i śródnabłonkowe
dowody anatomiczne wskazują na obecność 3 dróg śródnabłonkowych: (1) przedniego szlaku internodalnego, (2) środkowego przewodu intermodalnego i (3) tylnego przewodu intermodalnego.,
przednia droga intermodalna zaczyna się na przednim marginesie węzła SA i zakrzywia się przednio wokół SVC, aby wejść do przedniego pasma międzykomorowego, zwanego wiązką Bachmanna (patrz obrazek poniżej). Pasmo to kontynuuje do lewego przedsionka (LA), z przednim szlakiem intermodalnym wchodzącym do górnego marginesu węzła AV. Pęczek Bachmanna to duży pęczek mięśniowy, który wydaje się przewodzić impuls sercowy preferencyjnie z RZS do LA.
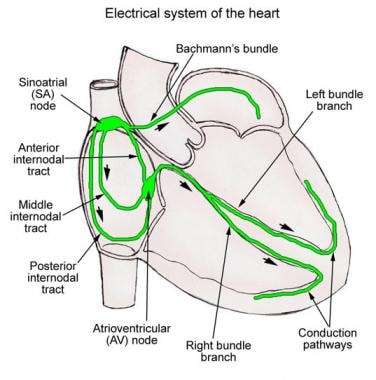 schematyczna ilustracja układu przewodzenia serca.,
schematyczna ilustracja układu przewodzenia serca., środkowy przewód międzykomorowy zaczyna się na Górnym i tylnym marginesie węzła zatokowego, przemieszcza się za SVC do grzebienia przegrody międzykomorowej i schodzi w przegrodzie międzykomorowej do górnego marginesu węzła AV.
tylny przewód międzykomorowy zaczyna się na tylnym brzegu węzła zatokowego i przemieszcza się tylnie wokół SVC i wzdłuż crista terminalis do grzbietu Eustachiusza, a następnie do przegrody międzykomorowej powyżej zatoki wieńcowej, gdzie łączy się z tylną częścią węzła AV., Te grupy tkanek intermodalnych są najlepiej określane jako intermodalne przedsionki mięśnia sercowego, a nie traktów, ponieważ nie wydają się być histologicznie dyskretne wyspecjalizowane traktów.
węzeł przedsionkowo-komorowy
zwarta część węzła przedsionkowo-komorowego (AV) jest powierzchowną strukturą znajdującą się tuż pod śródsierdziem RZ, przed ostium zatoki wieńcowej i bezpośrednio nad wkładką przegrody zastawki trójdzielnej., Znajduje się na wierzchołku trójkąta utworzonego przez pierścienia trójdzielnego i ścięgna Todaro, który pochodzi z centralnego ciała włóknistego i przechodzi tylnie przez przegrodę przedsionkową, aby kontynuować zastawkę Eustachiusza(patrz zdjęcia poniżej).
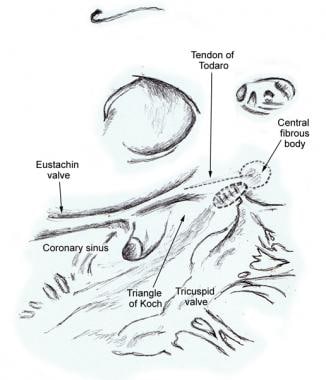 punktowany obszar przylegający do centralnego ciała włóknistego jest przybliżonym miejscem zwartego węzła przedsionkowo-komorowego. (Ilustracja oparta na Janse MJ, Anderson RH, McGuire MA, Ho SY. „AV węzłowy „reentry: Part I:” av węzłowy ” reentry revisited. J Cardiovasc Elektrofizjol. 1993 Oct;4 (5): 561-72.,)
punktowany obszar przylegający do centralnego ciała włóknistego jest przybliżonym miejscem zwartego węzła przedsionkowo-komorowego. (Ilustracja oparta na Janse MJ, Anderson RH, McGuire MA, Ho SY. „AV węzłowy „reentry: Part I:” av węzłowy ” reentry revisited. J Cardiovasc Elektrofizjol. 1993 Oct;4 (5): 561-72.,)  rysunek normalnego ludzkiego serca przedstawiający anatomiczne punkty orientacyjne trójkąta kocha. Trójkąt ten jest ograniczony przez ścięgno Todaro superiorly, włóknistą komis klapy chroniącej otwory żyły głównej dolnej i zatoki wieńcowej, przez mocowanie ulotki przegrody zastawki trójdzielnej inferiorly i przez usta zatoki wieńcowej u podstawy. (Ilustracja oparta na Janse MJ, Anderson RH, McGuire MA, Ho SY. „AV węzłowy „reentry: Part I:” av węzłowy ” reentry revisited. J Cardiovasc Elektrofizjol., 1993 Oct;4 (5): 561-72.)
rysunek normalnego ludzkiego serca przedstawiający anatomiczne punkty orientacyjne trójkąta kocha. Trójkąt ten jest ograniczony przez ścięgno Todaro superiorly, włóknistą komis klapy chroniącej otwory żyły głównej dolnej i zatoki wieńcowej, przez mocowanie ulotki przegrody zastawki trójdzielnej inferiorly i przez usta zatoki wieńcowej u podstawy. (Ilustracja oparta na Janse MJ, Anderson RH, McGuire MA, Ho SY. „AV węzłowy „reentry: Part I:” av węzłowy ” reentry revisited. J Cardiovasc Elektrofizjol., 1993 Oct;4 (5): 561-72.) w 85-90% ludzkich serc dopływ tętnicy do węzła AV jest gałęzią prawej tętnicy wieńcowej, która pochodzi z tylnego przecięcia AV i rowków międzykomorowych (crux). W pozostałych 10-15% serc gałąź lewej tętnicy wieńcowej obwodowej stanowi tętnicę węzłową AV. Włókna w dolnej części węzła AV mogą wykazywać automatyczne tworzenie impulsów. Główną funkcją węzła AV jest modulacja transmisji impulsu przedsionkowego do komór w celu koordynowania skurczów przedsionkowo-komorowych.,
pęczek His
pęczek His jest strukturą, która łączy się z dystalną częścią zwartego węzła AV, perforauje Centralne włókniste ciało i przechodzi przez pierścień włóknisty, gdzie nazywa się część niezwiązaną, ponieważ przenika przez membranous septum. Tkanka łączna centralnego ciała włóknistego i błoniastej przegrody zamyka penetrującą część wiązki AV, która może wysyłać rozszerzenia do centralnego ciała włóknistego., Komórki proksymalne części penetrującej są niejednorodne i przypominają komórki zwartego węzła AV; komórki dystalne są podobne do komórek w proksymalnych gałęziach pęczka.
gałęzie z przedniego i tylnego zstępującego tętnic wieńcowych zaopatrują górną przegrodę mięśniową międzykomorową w krew, co sprawia, że układ przewodzenia w tym miejscu jest bardziej odporny na uszkodzenie niedokrwienne, chyba że niedokrwienie jest rozległe.,
gałęzie pęczka Hisa
gałęzie pęczka Hisa powstają na górnym marginesie przegrody międzykomorowej mięśniowej, bezpośrednio poniżej przegrody błoniastej, a komórki lewej gałęzi pęczka Hisa kaskadują się w dół jako ciągły arkusz na przegrodę poniżej guzka aorty niekoronarnej. Prawa gałąź pęczka Hisa jest kontynuowana wewnątrzczaszkowo jako niezakłócone przedłużenie pęczka Hisa w dół prawą stroną przegrody międzykomorowej do wierzchołka prawej komory i podstawy mięśnia brodawkowatego przedniego., Anatomia systemu lewej odnogi pęczka Hisa może być zmienna i może nie odpowiadać stałemu podziałowi bifaskularnemu. Jednak do celów klinicznych i w elektrokardiografii (EKG) pojęcie trójglicerydów pozostaje użyteczne (patrz zdjęcia poniżej)
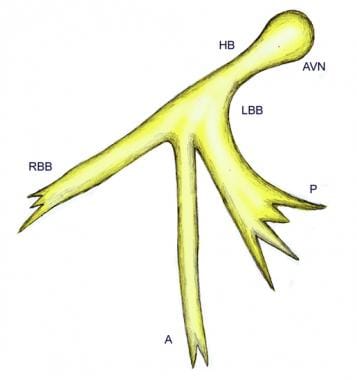 Schematyczne przedstawienie trójglicerydowych odgałęzień wiązki. A = przednia gałąź pęczka Hisa; AVN = węzeł przedsionkowo-komorowy; HB = pęczek Hisa; LBB = lewa gałąź pęczka Hisa; RBB = prawa gałąź pęczka Hisa; P = tylna gałąź pęczka Hisa.,
Schematyczne przedstawienie trójglicerydowych odgałęzień wiązki. A = przednia gałąź pęczka Hisa; AVN = węzeł przedsionkowo-komorowy; HB = pęczek Hisa; LBB = lewa gałąź pęczka Hisa; RBB = prawa gałąź pęczka Hisa; P = tylna gałąź pęczka Hisa., 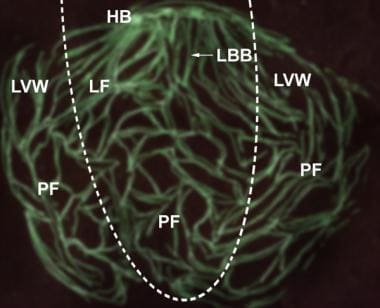 struktura systemu his-Purkinje w sercu myszy. Ekspresja Zielonego fluorescencyjnego białka była specjalnie ukierunkowana na komórki układu his-Purkinje u myszy. Pokazano zielone fluorescencyjne Sieci komórkowe w komorze lewej komory. Lewa ściana Komory (LVW) została wycięta od podstawy do wierzchołka, a następnie 2 części LVW zostały odciągnięte do tyłu, aby odsłonić lewą stronę przegrody międzykomorowej (LF). Linia przerywana wyznacza granicę między LF i LVW.,A = pęczek przedsionkowo-komorowy lewego pęczka; AVN = węzeł przedsionkowo-komorowy; HB = pęczek przedsionkowo-komorowy: LBB = gałąź lewej pęczka his; P = pęczek tylno-komorowy lewego pęczka his: RBB = gałąź prawej pęczki his: PF = włókno Purkinje. (Ilustracja oparta na Miquerol L, Meysen S, Mangoni m, et al. Asymetria architektoniczna i funkcjonalna układu His-Purkinje serca mysi. Cardiovasc Res. 2004 Jul 1; 63 (1): 77-86.,)
struktura systemu his-Purkinje w sercu myszy. Ekspresja Zielonego fluorescencyjnego białka była specjalnie ukierunkowana na komórki układu his-Purkinje u myszy. Pokazano zielone fluorescencyjne Sieci komórkowe w komorze lewej komory. Lewa ściana Komory (LVW) została wycięta od podstawy do wierzchołka, a następnie 2 części LVW zostały odciągnięte do tyłu, aby odsłonić lewą stronę przegrody międzykomorowej (LF). Linia przerywana wyznacza granicę między LF i LVW.,A = pęczek przedsionkowo-komorowy lewego pęczka; AVN = węzeł przedsionkowo-komorowy; HB = pęczek przedsionkowo-komorowy: LBB = gałąź lewej pęczka his; P = pęczek tylno-komorowy lewego pęczka his: RBB = gałąź prawej pęczki his: PF = włókno Purkinje. (Ilustracja oparta na Miquerol L, Meysen S, Mangoni m, et al. Asymetria architektoniczna i funkcjonalna układu His-Purkinje serca mysi. Cardiovasc Res. 2004 Jul 1; 63 (1): 77-86.,) końcowe włókna Purkinje
końcowe włókna Purkinje łączą się z końcami gałęzi pęczka, tworząc przeplatające się sieci na powierzchni wsierdzia obu komór, które przenoszą impuls serca prawie jednocześnie do całego serca prawej i lewej komory. Włókna Purkinjego są zwykle mniej skoncentrowane u podstawy komory i brodawkowatych końcówek mięśni. Przenikają tylko wewnętrzną trzecią endokardię. Włókna Purkinjego wydają się być bardziej odporne na niedokrwienie niż zwykłe włókna mięśnia sercowego.,
unerwienie węzła AV, jego pęczka i mięśnia sercowego
węzeł AV i jego pęczek są unerwione przez bogatą podaż włókien cholinergicznych i adrenergicznych o większej gęstości w porównaniu z mięśniem komorowym. Nerwy przywspółczulne do regionu węzła AV wchodzą do serca na skrzyżowaniu IVC i dolnej części LA, w sąsiedztwie zatoki wieńcowej ostium.,
autonomiczne wejście nerwowe do serca wykazuje pewien stopień „boczności”, z prawymi nerwami współczulnymi i błędnymi wpływającymi na węzeł SA bardziej niż węzeł AV i lewymi nerwami współczulnymi i błędnymi wpływającymi na węzeł AV bardziej niż węzeł SA. Dystrybucja wejścia neuronowego do węzłów SA i AV jest złożona ze względu na znaczne nakładające się unerwienie.,
Stymulacja prawego zwoju gwiaździstego powoduje częstoskurcz zatokowy z mniejszym wpływem na przewodzenie węzłowe AV, podczas gdy stymulacja lewego zwoju gwiaździstego zazwyczaj powoduje przesunięcie w rozruszniku zatokowym do miejsca pozamacicznego i konsekwentnie skraca czas przewodzenia węzłowego AV i refrakcję, ale niespójnie przyspiesza szybkość rozładowania węzła SA. Jednak stymulacja prawego nerwu błędnego szyjki macicy spowalnia szybkość rozładowania węzła SA, a stymulacja lewego nerwu błędnego przede wszystkim wydłuża czas przewodzenia węzłowego AV i refrakcję, gdy występuje boczność., Ani stymulacja współczulna, ani nerwowa nie wpływa na prawidłowe przewodzenie w wiązce.