| długotrwała potencja |
|
długotrwałe wzmacnianie (LTP) jest procesem, w którym wzmacniane są synapsy. Był przedmiotem wielu badań, ze względu na jego prawdopodobną rolę w kilku rodzajach pamięci. LTP jest przeciwieństwem depresji długoterminowej (LTD). W LTP, po intensywnej stymulacji neuronu presynaptycznego, zwiększa się amplituda odpowiedzi neuronu postsynaptycznego., Stosowany bodziec ma zazwyczaj krótki czas trwania (mniej niż 1 sekunda), ale wysoką częstotliwość (ponad 100 Hz). W neuronie postsynaptycznym bodziec ten powoduje wystarczającą depolaryzację do ewakuacji jonów magnezu, które blokują receptor NMDA, dzięki czemu duża liczba jonów wapnia dostaje się do dendrytu.
te jony wapnia są niezwykle ważnymi wewnątrzkomórkowymi przekaźnikami, które aktywują wiele enzymów zmieniając ich konformację. Jednym z tych enzymów jest kalmodulina, która staje się aktywna, gdy wiążą się z nią cztery jony wapnia., Następnie staje się Ca2+ / kalmodulina, głównym drugim posłańcem LTP. Następnie Ca2+ / kalmodulina aktywuje inne enzymy, które odgrywają kluczową rolę w tym procesie, takie jak cyklaza adenylanowa I II kinaza białkowa zależna od Ca2+/kalmoduliny (kinaza CaM II). Enzymy te z kolei modyfikują konformację przestrzenną innych cząsteczek, zwykle dodając do nich jon fosforanowy. Ten wspólny proces katalityczny nazywa się fosforylacją.,
|
w ten sposób aktywowana cyklaza adenylowa wytwarza cykliczny mono-fosforan adenozyny (cAMP), który z kolei katalizuje aktywność innego białka, kinazy a (lub PKA). Innymi słowy, istnieje typowa kaskada reakcji biochemicznych, które mogą mieć wiele różnych efektów. na przykład PKA fosforyluje receptory AMPA, dzięki czemu pozostają otwarte dłużej po związaniu z nimi glutaminianu. W rezultacie neuron postsynaptyczny ulega dalszej depolaryzacji, przyczyniając się w ten sposób do LTP., inne eksperymenty wykazały, że białko CREB jest kolejnym celem PKA. CREB odgrywa ważną rolę w transkrypcji genów, a jego aktywacja prowadzi do tworzenia nowych receptorów AMPA, które mogą jeszcze bardziej zwiększyć wydajność synaptyczną. inny enzym aktywowany przez Ca2+ / kalmodulinę, kinaza CaM II, ma właściwość decydującą o trwałości LTP: może się fosforylować! Jego aktywność enzymatyczna utrzymuje się długo po usunięciu wapnia z komórki i dezaktywacji Ca2+ / kalmoduliny., kinaza CaM II może z kolei fosforylować receptory AMPA i prawdopodobnie inne białka, takie jak kinazy MAP, które biorą udział w budowie dendrytów, lub same receptory NMDA, których przewodność wapnia zostanie zwiększona przez tę fosforylację. |
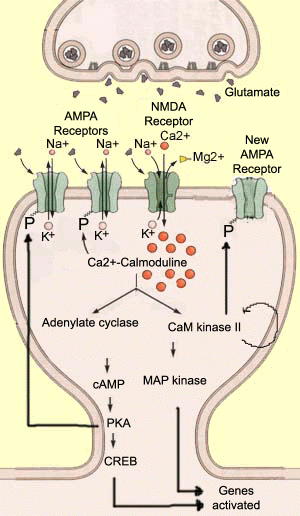 |
aby dać pewne wyobrażenie o złożoności sekwencji metabolicznych odpowiedzialny za LTP, wymienimy trzy inne enzymy obecnie badane., Kinaza białkowa C (PKC) wydaje się fosforylować receptory AMPA w tym samym miejscu co kinaza CaM II. Inhibitor 1 (ou I1) wydaje się być aktywowany przez PKA i zapobiega defosforylowaniu receptorów AMPA przez fosfatazę 1. Kinaza TYROZYNOWA SRC może być aktywowana bezpośrednio przez receptory AMPA, a następnie fosforylowana przez receptory NMDA.,
|
oprócz wszystkich mechanizmów postsynaptycznych biorących udział w tworzeniu LTP, od dawna postulowano, że pewne modyfikacje presynaptyczne występują podczas następującej po nich fazy podtrzymującej. Ale pewne modyfikacje, takie jak zwiększenie ilości glutaminianu uwalnianego przez neuron presynaptyczny, oznaczałyby obecność wstecznego przekaźnika, który wraca do tego neuronu i go modyfikuje., Ponieważ tlenek azotu (NO) jest gazem w stanie naturalnym i może w ten sposób rozpraszać się przez błony komórkowe, byłby idealnym kandydatem do tej roli. Ale jego zaangażowanie jest nadal przedmiotem wielu dyskusji i kontrowersji.