| POTENTIALISATION à LONG TERME |
|
la potentialisation à Long terme (LTP) est un processus dans lequel les synapses sont renforcées. Il a fait l’objet de nombreuses recherches, en raison de son rôle probable dans plusieurs types de mémoire. La LTP est le contraire de la dépression à long terme (LTD). En LTP, après une stimulation intense du neurone présynaptique, l’amplitude de la réponse du neurone post-synaptique augmente., Le stimulus appliqué est généralement de courte durée (moins de 1 seconde) mais de haute fréquence (plus de 100 Hz). Dans le neurone postsynaptique, ce stimulus provoque une dépolarisation suffisante pour évacuer les ions magnésium qui bloquent le récepteur NMDA, permettant ainsi à un grand nombre d’ions calcium d’entrer dans la dendrite.
ces ions calcium sont des messagers intracellulaires extrêmement importants qui activent de nombreuses enzymes en modifiant leur conformation. L’une de ces enzymes est la calmoduline, qui devient active lorsque quatre ions calcium s’y lient., Il devient alors Ca2 + / calmoduline, le second messager principal de la LTP. La Ca2 + / calmoduline active ensuite à son tour d’autres enzymes qui jouent un rôle clé dans ce processus, telles que l’adénylate cyclase et la protéine kinase II dépendante de la Ca2+/calmoduline (CaM kinase II). Ces enzymes modifient à leur tour la conformation spatiale d’autres molécules, généralement en leur ajoutant un ion phosphate. Ce processus catalytique commun est appelé phosphorylation.,
|
ainsi, l’adénylate cyclase activée produit de l’adénosine monophosphate cyclique (AMPc), qui à son tour catalyse l’activité d’une autre protéine, la kinase A (ou PKA). En d’autres termes, il existe une cascade typique de réactions biochimiques qui peuvent avoir de nombreux effets différents. Par exemple, le PKA phosphoryle les récepteurs AMPA, ce qui leur permet de rester ouverts plus longtemps après que le glutamate s’y lie. En conséquence, le neurone postsynaptique se dépolarise davantage, contribuant ainsi à la LTP., D’autres expériences ont montré que la protéine CREB est une autre cible du PKA. CREB joue un rôle majeur dans la transcription des gènes, et son activation conduit à la création de nouveaux récepteurs AMPA qui peuvent augmenter encore l’efficacité synaptique. l’autre enzyme activée par la Ca2+/calmoduline, la CaM kinase II, possède une propriété décisive pour la persistance de la LTP: elle peut se phosphoryler! Son activité enzymatique se poursuit longtemps après l’évacuation du calcium de la cellule et la désactivation du Ca2+/calmoduline., La CaM kinase II peut alors à son tour phosphoryler les récepteurs AMPA et probablement d’autres protéines telles que les MAP kinases, qui sont impliquées dans la construction des dendrites, ou les récepteurs NMDA eux-mêmes, dont la conductance calcique serait augmentée par cette phosphorylation. |
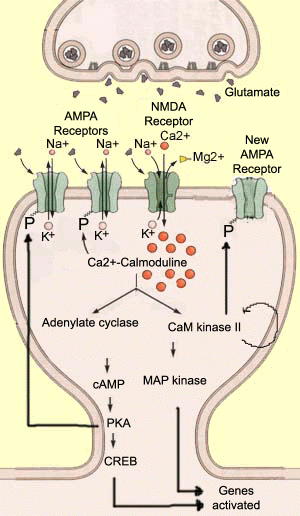 |
Pour donner une idée de la complexité des séquences métaboliques responsables de la LTP, nous allons en mentionner trois autres enzymes actuellement à l’étude., La protéine kinase C (PKC) semble phosphoryler les récepteurs AMPA au même site que la CaM kinase II. L’inhibiteur 1 (ou I1) semble être activé par la PKA et empêche la phosphatase 1 de déphosphoryler les récepteurs AMPA. Et la tyrosine kinase SRC peut être activée directement par les récepteurs AMPA, puis phosphoryler les récepteurs NMDA.,
|
en plus de tous les mécanismes post-synaptiques impliqués dans l’établissement de LTP, il a longtemps été postulé que certaines modifications présynaptiques se produisent pendant la phase de maintenance qui a suivi. Mais certaines modifications, telles qu’une augmentation de la quantité de glutamate libéré par le neurone présynaptique, impliquerait la présence d’un messager rétrograde qui remonte à ce neurone et la modifie., Parce que l’oxyde nitrique (NO) est un gaz à l’état naturel, et peut ainsi diffuser à travers les membranes cellulaires, il serait un candidat idéal pour ce rôle. Mais son implication fait encore l’objet de nombreux débats et controverses.