| DLOUHODOBÉ POTENCIACE |
|
Long-term potenciace (LTP) je proces, ve kterém jsou synapse posíleny. To bylo předmětem velkého výzkumu, protože jeho pravděpodobná role v několika typech paměti. LTP je opakem dlouhodobé deprese (LTD). V LTP se po intenzivní stimulaci presynaptického neuronu zvyšuje amplituda reakce postsynaptického neuronu., Aplikovaný stimul má obvykle krátké trvání (méně než 1 sekundu), ale vysokou frekvenci (přes 100 Hz). V postsynaptickém neuronu, tento podnět způsobí dostatečné depolarizace evakuovat hořečnatých iontů, které blokují NMDA receptoru, což umožňuje velké množství iontů vápníku vstoupit dendrit.
tyto ionty vápníku jsou nesmírně důležitými intracelulárními posly, kteří aktivují mnoho enzymů změnou jejich konformace. Jedním z těchto enzymů je kalmodulin, který se stává aktivním, když se na něj váží čtyři ionty vápníku., Poté se stává Ca2+ / calmodulin, hlavní druhý posel pro LTP. Ca2+/kalmodulin pak zase aktivuje další enzymy, které hrají klíčovou úlohu v tomto procesu, jako jsou adenylát cyklázy a Ca2+/kalmodulin-dependentní proteinkinázy II (CaM kináza II). Tyto enzymy zase modifikují prostorovou konformaci jiných molekul, obvykle přidáním fosfátového iontu k nim. Tento společný katalytický proces se nazývá fosforylace.,
|
Tak, aktivovaný adenylátcyklázu vyrábí cyklický adenosin mono-vápenatý (cAMP), což katalyzuje činností jiné proteiny, kinázy A (nebo PKA). Jinými slovy, existuje typická kaskáda biochemických reakcí, které mohou mít mnoho různých účinků. například pKa fosforyluje AMPA receptory, což jim umožňuje zůstat otevřené déle poté, co se na ně glutamát váže. V důsledku toho se postsynaptický neuron dále depolarizuje, čímž přispívá k LTP., další experimenty ukázaly, že CREB protein je dalším cílem PKA. CREB hraje hlavní roli v genové transkripci a jeho aktivace vede k vytvoření nových AMPA receptorů, které mohou ještě dále zvyšovat synaptickou účinnost. druhý enzym aktivovaný Ca2+ / calmodulinem, CaM kinase II, má vlastnost, která je rozhodující pro přetrvávání LTP: může se fosforylovat sám! Jeho enzymatická aktivita pokračuje dlouho poté, co byl vápník evakuován z buňky a Ca2+/calmodulin byl deaktivován., CaM kinázy II může následně fosforylují receptory AMPA, a pravděpodobně i další proteiny, jako jsou MAP kinázy, které jsou zapojeny v budově dendrity, nebo NMDA receptory samy, jehož vodivost vápník by být zvýšena tím, že tato fosforylace. |
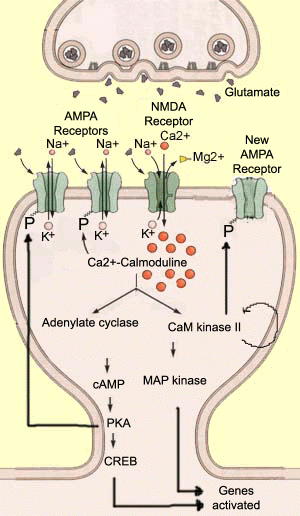 |
dát nějakou představu o složitosti metabolických sekvencí zodpovědné za LTP, zmíníme tři další enzymy v současné době zkoumán., Protein kinázy C (PKC) se zdá, že fosforylují AMPA receptory na stejném místě jako CaM kinázy II. Inhibitor 1 (ou I1) se zdá být aktivuje PKA a zabránit fosfatázy 1 z dephosphorylating AMPA receptory. A TYROSINKINÁZA SRC může být aktivována přímo receptory AMPA a poté fosforylovat receptory NMDA.,
|
kromě všech post-synaptické mechanismy podílející se na vzniku LTP, to dlouho bylo předpokládal, že některé presynaptické změny vyskytují během následné udržovací fáze. Některé modifikace, jako je zvýšení množství glutamátu uvolněného presynaptickým neuronem, by však znamenaly přítomnost retrográdního posla, který se vrací k tomuto neuronu a modifikuje ho., Protože oxid dusnatý (ne) je plyn ve svém přirozeném stavu a může se tak rozptýlit buněčnými membránami, byl by ideálním kandidátem na tuto roli. Jeho zapojení je však stále předmětem mnoha debat a kontroverzí.